Les plastes peuvent être différents : types, structure, fonctions. Qu'est-ce qu'un plaste en biologie ? En combien de types les plastes sont-ils divisés ?
Les plastes sont des organites protoplastes, caractéristiques uniquement des cellules végétales. Seules les bactéries, les algues bleu-vert et éventuellement les champignons n’en possèdent pas.
Chez les plantes supérieures, les plastes se trouvent dans les cellules végétatives adultes de tous les organes - dans la tige, la feuille, la racine et la fleur. Les plastes sont des organites relativement gros, beaucoup plus gros que les mitochondries, et parfois même plus gros que le noyau, plus denses que le cytoplasme environnant, clairement visibles au microscope optique. Ils ont une structure caractéristique et remplissent diverses fonctions, principalement liées à la synthèse de substances organiques.
Dans une cellule végétale adulte, selon la couleur, la forme et la fonction, on distingue trois principaux types de plastes : les chloroplastes (plastes verts), les chromoplastes (plastes jaunes et oranges) et les leucoplastes (plastes incolores). Ces derniers sont de plus petite taille que les deux types de plastes précédents.
Chloroplastes
La base structurelle du chloroplaste est constituée de protéines (environ 50 % du poids sec), elles contiennent également 5 à 10 % de chlorophylle et 1 à 2 % de caroténoïdes. Comme dans les mitochondries, une petite quantité d'ARN (0,5 à 3,5 %) et encore moins d'ADN ont été trouvées dans les chloroplastes. L'importance exceptionnelle des chloroplastes réside dans le fait que le processus de photosynthèse s'y déroule. L'amidon produit lors de la photosynthèse est dit primaire, ou assimilatif, et se dépose dans les chloroplastes sous forme de petits grains d'amidon. Pour une photosynthèse normale, la présence de chlorophylle est nécessaire. La chlorophylle est le principal principe actif de la photosynthèse. Il absorbe l’énergie lumineuse et la dirige pour effectuer des réactions photosynthétiques. La chlorophylle peut être extraite des plastes à l'aide d'alcool, d'acétone ou d'autres solvants organiques. Le rôle des pigments jaunes dans la photosynthèse n’est pas encore bien compris. On suppose qu'ils absorbent également l'énergie solaire et la transfèrent à la chlorophylle, ou qu'ils effectuent avec elle des réactions spécifiques importantes pour la photosynthèse.
Conformément à leurs fonctions, les chloroplastes se trouvent principalement dans les organes et tissus photosynthétiques exposés à la lumière - dans les feuilles et les jeunes tiges, les fruits non mûrs. Parfois, les chloroplastes se trouvent même dans les racines, par exemple dans les racines adventives du maïs. Mais leur quantité principale est concentrée dans les cellules mésophylles (pulpe) de la feuille.
Contrairement aux autres organites, les chloroplastes des plantes supérieures se caractérisent par l'uniformité et la constance de leur forme et de leur taille. Le plus souvent, ils ont une forme en forme de disque ou de lentille et, lorsqu'ils sont posés à plat, ils ont un contour rond ou polygonal. Dans ce cas, ils sont souvent aussi appelés grains de chlorophylle. La taille des chloroplastes est assez constante et même chez différentes espèces de plantes supérieures, elle varie dans des limites insignifiantes, en moyenne de 3 à 7 microns (épaisseur de 1 à 3 microns). Les chloroplastes plus gros sont rares dans les plantes supérieures. Par exemple, chez Selaginella (lycophytes), on trouve un ou deux grands chloroplastes lamellaires dans les cellules de la peau des feuilles. La taille et la forme des chloroplastes changent en fonction des conditions extérieures. Chez les plantes qui aiment l'ombre, les chloroplastes sont généralement plus gros que chez les plantes qui aiment la lumière et, en règle générale, sont plus riches en chlorophylle. Généralement, une cellule porte un grand nombre de chloroplastes et leur nombre varie considérablement ; en moyenne, il contient de 20 à 50 chloroplastes. Les feuilles, ainsi que les jeunes fruits non mûrs, sont particulièrement riches en chloroplastes. Le nombre total de chloroplastes dans une plante peut être énorme ; par exemple, dans un arbre adulte, il y a des dizaines et des centaines de milliards de chloroplastes. Le nombre de chloroplastes dans une cellule est lié à leur taille. Ainsi, dans le maïs, les cellules des feuilles contiennent généralement plusieurs chloroplastes, mais dans les variétés à chloroplastes particulièrement gros, leur nombre par cellule est réduit à deux.
Dans de nombreuses plantes inférieures (algues), la forme, le nombre et la taille des chloroplastes sont très divers. Ils peuvent être en forme de plaque (Mougeotia), d'étoile (Zygnema), ou encore sous forme de rubans spiralés (Spirogyra) et de cylindres nervurés (Closterium). Ces chloroplastes sont généralement très gros, présents en petit nombre dans la cellule (de un à plusieurs) et sont appelés chromatophores. Mais les algues peuvent également avoir des chloroplastes de forme lenticulaire habituelle, et dans ce cas, leur nombre dans la cellule est généralement important.
Dans les cellules des plantes supérieures, les chloroplastes sont situés dans le cytoplasme de telle sorte qu'un de leurs côtés plats fait face à la membrane cellulaire, et ils sont surtout nombreux à proximité des espaces intercellulaires remplis d'air. Ici, ils sont étroitement adjacents les uns aux autres et leurs contours deviennent anguleux. Cependant, la position des chloroplastes dans la cellule peut changer en fonction des conditions extérieures et surtout de la lumière. Ils sont situés dans la cage de manière à capter la lumière de la meilleure façon possible, sans être exposés directement au soleil. Dans les feuilles de certaines plantes, sous une lumière diffuse, les chloroplastes sont situés principalement sur les parois de la membrane cellulaire qui font face à la surface de l'organe, sous une lumière vive, ils se concentrent sur les parois latérales ou se tournent vers les rayons avec un côté étroit ; , c'est-à-dire avec un bord. Le même mouvement des chloroplastes est parfois observé sous l'influence d'autres stimuli - température, produits chimiques, mécaniques, etc. La question de savoir si le mouvement des plastes est actif ou passif (flux cytoplasmique) n'a pas encore été complètement clarifiée, mais il existe actuellement davantage d'arguments. en faveur du mouvement actif.
Sur la base de la complexité des processus de photosynthèse, constitués d'un certain nombre de réactions, chacune catalysée par une enzyme spécifique, on peut supposer que les chloroplastes ont une structure ordonnée et complexe. En effet, déjà au microscope optique ordinaire, il est souvent visible que les chloroplastes ne sont pas complètement homogènes, mais on y observe de petits grains plus foncés, orientés parallèlement à la surface du chloroplaste, appelés céréales. Des études au microscope électronique ont confirmé l'existence du grana et ont montré que l'ensemble du chloroplaste et du grana ont une structure complexe.
Comme les mitochondries, les chloroplastes sont des structures membranaires qui se trouvent librement dans le cytoplasme. Ils sont séparés du cytoplasme par une membrane à deux membranes avec un espace lumineux clairement visible entre les membranes. Ces membranes sont supposées lisses et exemptes de particules attachées. Jusqu'à récemment, on croyait que la membrane chloroplastique était solide, dépourvue de trous et que ses membranes n'étaient pas reliées aux membranes du réticulum endoplasmique. Mais les preuves s’accumulent désormais pour montrer que ce n’est pas toujours le cas. Parfois, des trous submicroscopiques peuvent apparaître à certains endroits de la coquille. Il est possible que pendant certaines périodes d'activité chloroplastique ces « pores » soient en contact étroit avec le réticulum endoplasmique, mais ce contact est de courte durée. L'enveloppe chloroplastique, ayant la propriété de perméabilité sélective, joue un rôle régulateur dans l'échange de substances entre le cytoplasme et le chloroplaste.
Le corps du chloroplaste est pénétré par un système de plaques à double membrane appelées lamelles. L'espace entre les lamelles est rempli d'un fluide protéique aqueux - stroma, ou la matrice chloroplastique. Le stroma peut contenir des grains d'amidon, des gouttelettes d'huile et des particules ressemblant à des ribosomes. Tout récemment, grâce à des méthodes de préparation particulièrement sophistiquées, des amas de fibrilles parallèles d'un diamètre de 80 à 100 Å et d'une longueur supérieure à 1 000 Å ont été découverts dans le stroma des chloroplastes de certaines plantes. Ces faisceaux de microfibrilles chloroplastiques sont appelés stromacentres. Leur fonction n'est absolument pas claire.
Dans certaines zones du chloroplaste, les lamelles s'emboîtent assez étroitement les unes aux autres, situées parallèlement à sa surface, ce qui entraîne la formation d'amas de lamelles appelés grana dans ces zones. À l'intérieur du grana, les membranes appariées des lamelles fusionnent sur les bords, formant des sacs aplatis fermés appelés disques, ou thylakoïdes. Les paquets de ces disques forment un grana. Les grains individuels sont interconnectés en un seul système à l'aide de lamelles qui pénètrent dans les espaces intergranulaires. La chlorophylle n'est pas dispersée de manière diffuse dans tout le chloroplaste, mais est concentrée dans les lamelles, vraisemblablement sous la forme d'une couche monomoléculaire. Les ribosomes se trouvent non seulement dans la matrice, mais également à la surface du grana.

Le nombre de disques en grains varie de deux à plusieurs dizaines et le diamètre, selon le type de plante, de 0,3 à 2 microns. Par conséquent, chez de nombreuses plantes, les grana ne sont pas visibles au microscope optique. Le nombre et l'emplacement des grana dans un chloroplaste dépendent du type de plante, de l'âge et de l'activité des chloroplastes. Il y a tellement de granules dans les chloroplastes d'Aspidistra qu'ils se touchent, et dans les cellules satellites de citrouille, le volume principal du chloroplaste est occupé par le stroma. Dans les chloroplastes des feuilles de tomate et de chrysanthème, les grana sont dispersés de manière aléatoire et dans les chloroplastes du tabac, ils sont correctement orientés par rapport à la surface du chloroplaste et sont situés à égale distance les uns des autres. Les plantes qui aiment la lumière ont des grains plus petits que les plantes tolérantes à l'ombre.
La structure des chloroplastes des plantes supérieures est parfaitement adaptée pour remplir leur fonction principale : la photosynthèse. La division même de l'appareil porteur de chlorophylle en petits plastes entraîne une énorme augmentation de la surface active. En raison de la formation de membranes et de grana, cette surface augmente encore plus. La grande surface active et l’orientation spatiale fine permettent un accès facile à l’énergie quantique lumineuse et la capacité de transférer cette énergie aux systèmes chimiques impliqués dans la photosynthèse. Le principe des chambres fermées - les thylakoïdes, en raison de la séparation spatiale, permet d'effectuer simultanément et indépendamment le même complexe de réactions qui composent la photosynthèse. La synthèse des protéines se produit dans les ribosomes des chloroplastes.
Dans les cellules de certaines algues (spirogyres) et plus rarement de plantes supérieures (par exemple, les cellules de ce qu'on appelle la gaine des faisceaux vasculaires du maïs), il existe des chloroplastes sans visage dans lesquels les lamelles pénètrent dans le stroma sans former de granae distincts.
L'origine et le développement des chloroplastes ont été très peu étudiés et il n'y a pas encore de point de vue unique sur cette question. On sait que dans les jeunes cellules embryonnaires, il n'y a pas de chloroplastes différenciés. Au lieu de cela, il existe ce qu'on appelle proplastes. Ce sont des corps très petits (fractions d’un micron), situés à la limite de la résolution d’un microscope optique. Initialement, ils ont une forme amiboïde (lobes d'ours), sont délimités du cytoplasme par une double membrane et ne contiennent ni membranes internes ni chlorophylle. Les membranes internes qui forment les lamelles se développent plus tard. Il existe plusieurs hypothèses sur le développement ultérieur des proplastes. Selon l'un d'eux, dans le stroma transparent du proplastide, se forment d'abord des amas de minuscules vésicules, disposées dans le bon ordre comme un réseau cristallin. Cette accumulation de bulles, dont chacune est recouverte de sa propre membrane, est appelée facette primaire. Des lamelles apparaissent à la périphérie du grana primaire, qui s'étendent dans toutes les directions à partir de celui-ci. Par la suite, toutes les structures lamellaires du chloroplaste, y compris le grana, se forment dessus. Lorsqu'ils sont exposés à la lumière, ils présentent des dépôts de pigments et surtout de chlorophylle.
Selon une autre hypothèse, les lamelles seraient initialement formées de plis de la membrane interne de la coque du proplastide, et non de vésicules. Dans ce cas, une structure semblable à une mitochondrie apparaît pour la première fois.
Ainsi, ces hypothèses reposent sur le principe de continuité des plastes et nient leur origine à partir d’autres organites du protoplaste et surtout du cytoplasme. Cependant, d’autres scientifiques pensent que les mitochondries et les plastes sont d’origine étroitement liée. Par exemple, il a été possible de démontrer l’émergence de mitochondries à partir de chloroplastes matures par « bourgeonnement ». Par la suite, ces mitochondries pourraient à nouveau s’unir aux chloroplastes. Mais toutes ces hypothèses n'ont pas encore été suffisamment étayées, et la question de l'origine des plastes attend toujours sa solution.
En plus d'être issus des proplastes, les chloroplastes peuvent se reproduire par simple division. Dans ce cas, deux plastes filles sont formés à partir du chloroplaste adulte, souvent de tailles inégales. L’image au microscope électronique d’une telle division n’a pas encore été étudiée.
La structure du chloroplaste ne reste pas constante ; elle change naturellement au cours de la croissance cellulaire. Le changement dans la structure des chloroplastes avec l'âge des feuilles est perceptible même au microscope optique. Ainsi, les jeunes feuilles ont généralement une structure à grain fin, tandis que les feuilles d'âge moyen ont une structure à grain grossier. Dans les feuilles vieillissantes, la structure et la dégradation des chloroplastes se produisent.
Les chloroplastes sont des organites plutôt délicats et fragiles. Lorsque les cellules sont placées dans de l'eau distillée ou une solution saline hypotonique, elles gonflent rapidement, des gonflements semblables à des bulles se forment à leur surface, puis elles se brouillent. Des études en microscopie électronique ont montré que le gonflement se produit dans le stroma et non dans les lamelles. Lorsqu'une cellule est endommagée, les chloroplastes, observés au microscope conventionnel, deviennent d'abord grossièrement granuleux, gonflent, prennent un aspect mousseux et finalement la granularité disparaît. Des changements pathologiques se produisent dans les chloroplastes des feuilles et en cas de manque d'éléments nutritifs minéraux dans le sol. Cependant, les chloroplastes de certaines cellules peuvent également présenter une grande résistance. Ainsi, chez les arbres, la couleur verte de l'écorce est due à la présence d'une couche de cellules comportant des chloroplastes. Ces chloroplastes tolèrent bien les basses températures et entrent dans un état actif, qui se détecte par le fort verdissement de l'écorce, par exemple chez le tremble, très tôt au printemps, alors qu'il y a encore de fortes gelées la nuit. Les basses températures hivernales sont également tolérées par les chloroplastes des feuilles (aiguilles) de nos conifères à feuilles persistantes. Dans le même temps, comme l’ont montré des études au microscope électronique, ils conservent leur organisation interne complexe.
Leucoplastes
Ce sont de petits plastes incolores. Ils sont souvent difficiles à détecter au microscope optique car ils sont incolores et ont le même indice de réfraction que le cytoplasme. Ils ne peuvent être détectés que si de grosses inclusions s'accumulent à l'intérieur. Ce sont des organites très délicats et, lors de la préparation de sections de matière vivante, ils sont détruits encore plus facilement que les chloroplastes. On les retrouve dans les cellules adultes cachées du soleil : dans les racines, les rhizomes, les tubercules (pommes de terre), les graines, les noyaux de tiges, mais aussi dans les cellules exposées à une forte lumière (cellules de la peau). Les leucoplastes se rassemblent souvent autour du noyau, l'entourant parfois de tous côtés. La forme des leucoplastes est très variable, il s'agit le plus souvent de formations sphériques, ovoïdes ou fusiformes.

Les leucoplastes sont des organites associés à la formation de nutriments de réserve - amidon, protéines et graisses. L'activité des leucoplastes est spécialisée : certains d'entre eux accumulent principalement de l'amidon (amyloplastes), d'autres - des protéines (protéoplastes, également appelés aleuronoplastes), et d'autres - des huiles (oléoplastes). Les leucoplastes des cellules cutanées des feuilles et des tiges ne peuvent pas être classées dans l'un de ces types, car leur fonction n'a pas encore été clarifiée.
Amyloplastes accumuler de l'amidon sous forme de soi-disant grains d'amidon. C'est le type prédominant de leucoplaste. La structure des amyloplastes et le mécanisme de formation de l'amidon sont difficiles à étudier au microscope optique, et encore peu étudiés au microscope électronique. On suppose qu'ils sont formés à partir de proplastes, mais contrairement aux chloroplastes, le développement de leur structure ne va pas loin, mais est retardé à un stade assez précoce - le stade d'un plaste immature et faiblement lamellaire. A l'extérieur, les amyloplastes sont délimités par une membrane à double membrane. L'intérieur du plaste est rempli d'un stroma finement granuleux. La formation de grains d'amidon dans les amyloplastes est précédée du développement de minuscules vésicules qui fusionnent et s'aplatissent, limitant la zone stromale au centre du plaste avec une membrane. Cette zone, appelée centre éducatif, devient plus léger, ressemblant à une vacuole. Le dépôt d'amidon commence dans le centre éducatif. Lorsque le futur grain d'amidon commence à grossir, les membranes délimitant le centre éducatif disparaissent et la croissance du grain se produit sans lien avec elles. Lorsque l'amidon est déposé, la coquille et le stroma de l'amyloplaste peuvent être considérablement étirés, ce qui entraîne une augmentation importante de la taille de l'amyloplaste en raison de la croissance du grain d'amidon. Le grain d’amidon remplit alors la quasi-totalité de la cavité de l’amyloplaste, repoussant son contenu vivant vers la périphérie sous la forme d’une fine pellicule à la surface du grain. Dans de nombreux cas, le grain d’amidon peut atteindre une taille telle que l’amyloplaste se rompt et ne reste que sur un côté du grain d’amidon. Dans ce cas, de nouvelles portions d'amidon ne peuvent être synthétisées que dans les zones où le grain d'amidon reste en contact avec les membranes et le stroma de l'amyloplaste.
Se développant à partir de proplastes, les amyloplastes, dans certaines conditions, peuvent se transformer en plastes d'autres types. Si, par exemple, vous placez une racine d'orge à la lumière, vous remarquerez que certains leucoplastes augmentent de volume et se transforment en chloroplastes, semblables à ceux qui se forment dans les feuilles. Si une telle racine est à nouveau privée de lumière, alors ces chloroplastes diminuent de taille et perdent leur chlorophylle, mais ne se transforment pas en leucoplastes, mais produisent des globules (boules) de carotène, devenant ainsi des chromoplastes. Oléoplastes, c'est-à-dire que les leucoplastes, qui forment principalement des huiles, se trouvent beaucoup moins fréquemment que les amyloplastes (par exemple dans les cellules des feuilles de certaines monocotylédones). Ils sont généralement le produit du vieillissement des chloroplastes qui perdent de la chlorophylle. Dans ce cas, de minuscules globules d’huile apparaissent dans le stroma du plaste. La coquille du plaste se décompose alors et le contenu des plastes adjacents fusionne, formant de plus grosses gouttelettes de graisse. Parfois, l'amidon s'accumule également dans ces plastes.
La synthèse des protéines de stockage - les protéines - peut être réalisée dans le troisième type de leucoplastes - protéoplastes. Les protéines sous forme de cristaux et de grains se forment dans les graines de nombreuses plantes, en particulier celles qui contiennent également beaucoup d'huile (par exemple, les graines de ricin). Les protéoplastes, comme les amyloplastes, proviennent des proplastes. Leur développement est également retardé au stade immature du plaste lamellaire. Dans le stroma du protéoplaste, la protéine de stockage s’accumule initialement sous forme de fibrilles, qui sont ensuite regroupées en faisceaux plus gros. Ensuite, la coque et le stroma du plaste sont détruits et des faisceaux de fibrilles protéiques se transforment en une sorte de petites vacuoles visqueuses. Ensuite, les vacuoles des plastes voisins fusionnent et une partie de la protéine se forme sous forme de cristalloïdes.
Ainsi, l’amidon, les protéines de stockage et les gouttelettes d’huile sont des inclusions inertes, des déchets des plastes. De plus, chacun d'entre eux peut s'accumuler non seulement dans les leucoplastes, mais également dans les chloroplastes et les chromoplastes. Mais si l'amidon est formé dans les plastes, alors les protéines et les graisses de stockage peuvent très souvent être d'origine extraplastidienne, probablement directement dans le cytoplasme et indépendamment des plastes. Les processus structurels se produisant dans ce cas sont encore peu étudiés.
Chromoplastes
Les chromoplastes sont des plastes jaunes ou oranges et même rouges. On les retrouve dans les cellules de nombreux pétales (pissenlit, renoncule, souci), de fruits mûrs (tomates, églantier, sorbier des oiseleurs, citrouille, pastèque, orange) et de légumes-racines (carottes, betteraves fourragères). La couleur vive de ces organes est due aux pigments jaunes et oranges - les caroténoïdes, concentrés dans les chromoplastes. Ces pigments sont également caractéristiques des chloroplastes, mais là ils sont masqués par la chlorophylle. Ils sont insolubles dans l'eau, mais solubles dans les graisses.
Contrairement aux chloroplastes, la forme des chromoplastes est très variable et est déterminée par leur origine et l'état de leurs pigments, ainsi que par la position systématique de la plante qui les forme. Selon l'état des caroténoïdes, on distingue trois types de chromoplastes :
- les chromoplastes, dans lesquels les caroténoïdes se déposent sous la forme de petits cristaux sans rapport entre eux, visibles au microscope optique (chromoplastes de carotte) ;
- les chromoplastes, dans lesquels les caroténoïdes sont dissous dans des globules lipoïdes submicroscopiques (pétales de renoncule et d'aloès) ;
- les chromoplastes dont les caroténoïdes sont rassemblés en faisceaux constitués de fils submicroscopiques et associés à des fibrilles protéiques (poivrons rouges, tomates, mandarines).
Contrairement aux chloroplastes et aux leucoplastes, les chromoplastes proviennent rarement directement des proplastes, mais sont généralement le résultat d'une dégénérescence des chloroplastes. Une exception concerne les chromoplastes de carotte, qui ne proviennent pas de chloroplastes, mais de leucoplastes ou directement de proplastes. Les parties des racines qui ne sont pas immergées dans le sol et se développent à la lumière deviennent généralement vertes. Cela ne se produit pas à la suite de la transformation de chromoplastes en chloroplastes, mais à cause de la formation de chloroplastes à partir de proplastes ou de leucoplastes. Les chromoplastes ne peuvent pas du tout se transformer en d’autres types de plastes. Le plus souvent, les chromoplastes se forment lors de la destruction des chloroplastes, lorsque ces derniers entrent dans une phase irréversible de développement. C'est l'origine des chromoplastes de types 2 et 3. Dans le même temps, la teneur en graisses et en caroténoïdes des chloroplastes augmente, qui sont collectés dans le stroma du plaste sous forme de globules submicroscopiques, les structures lamellaires disparaissent et la chlorophylle est détruite. Les globules pigmentaires se développent et le volume du stroma diminue ; les globules peuvent donc remplir la majeure partie du plaste. La forme ronde du chloroplaste « mère » est préservée. Un processus similaire de dégradation des chloroplastes se produit probablement lors du jaunissement automnal des feuilles et lors de la maturation des fruits. La chlorophylle des feuilles jaunissantes est détruite et cesse de masquer les caroténoïdes, qui dépassent fortement et provoquent la couleur jaune des feuilles.
Dans les racines de carotte, les chromoplastes proviennent de leucoplastes, initialement féculents, tandis que les caroténoïdes s'accumulent dans le stroma du plaste, qui se cristallisent ensuite. L'amidon disparaît à mesure que la concentration de carotène augmente, et la masse des plastes diminue et devient difficile à détecter. Le pigment cristallisé constitue le volume prédominant du chromoplaste, donc la forme du chromoplaste est finalement déterminée par la forme du pigment cristallisé et est généralement irrégulière : dentelée, en forme de croissant, d'aiguille ou lamellaire. Les plaques peuvent avoir la forme d'un triangle, d'un losange, d'un parallélogramme, etc.

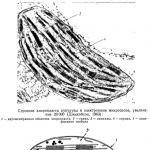
La photo montre l’une des cellules de pastèque avec de la pulpe de framboise vue au microscope optique. La cellule contient un cytoplasme visible, constitué de minces filaments étirés dans différentes directions. Dans les brins les plus massifs du cytoplasme se trouvent des cristaux en forme d'aiguilles de pigment chromoplaste. La plus grande accumulation de cristaux est observée près du noyau. Dans une autre variété de pastèque à chair carmin, le pigment chromoplaste cristallise non seulement sous forme de cristaux en forme d'aiguilles, mais également de prismes courts de différentes tailles.
L’importance des chromoplastes dans le métabolisme est très peu comprise. Comme les leucoplastes, ils n’ont pas la capacité de photosynthèse car ils ne contiennent pas de chlorophylle. L'importance indirecte des chromoplastes est qu'ils déterminent la couleur vive des fleurs et des fruits, ce qui attire les insectes pour la pollinisation croisée et d'autres animaux pour la distribution des fruits.
Si vous trouvez une erreur, veuillez surligner un morceau de texte et cliquer sur Ctrl+Entrée.
Les plastes sont des organites membranaires présents dans les organismes eucaryotes photosynthétiques (plantes supérieures, algues inférieures, certains organismes unicellulaires). Tout un ensemble de plastes différents (chloroplaste, leucoplaste, amyloplaste, chromoplaste) ont été trouvés dans les plantes supérieures, représentant une série de transformations mutuelles d'un type de plaste en un autre. La principale structure qui effectue les processus photosynthétiques est le chloroplaste (Fig. 226a).
Chloroplaste. Comme déjà indiqué, la structure du chloroplaste rappelle en principe la structure de la mitochondrie. Il s'agit généralement de structures allongées d'une largeur de 2 à 4 microns et d'une longueur de 5 à 10 microns. Les algues vertes possèdent des chloroplastes géants (chromatophores) atteignant une longueur de 50 microns. Le nombre de chloroplastes dans les cellules végétales varie. Ainsi, les algues vertes peuvent avoir un chloroplaste, les plantes supérieures en ont en moyenne 10 à 30, et dans les cellules géantes du tissu palissadique du cormoran, environ 1 000 chloroplastes par cellule ont été trouvés.
La membrane externe des chloroplastes, comme la membrane interne, a une épaisseur d'environ 7 microns ; ils sont séparés les uns des autres par un espace intermembranaire d'environ 20 à 30 nm. La membrane interne des chloroplastes sépare le stroma plastidien, qui est similaire à la matrice mitochondriale. Dans le stroma du chloroplaste mature des plantes supérieures, deux types de membranes internes sont visibles. Ce sont des membranes qui forment des lamelles stromales plates et étendues et des membranes de thylakoïdes, des vacuoles ou des sacs plats en forme de disque.
Les lamelles stromales (environ 20 µm d'épaisseur) sont des sacs creux plats ou ont l'apparence d'un réseau de canaux ramifiés et interconnectés situés dans un même plan. En règle générale, les lamelles stromales à l'intérieur du chloroplaste sont parallèles les unes aux autres et ne forment pas de connexions entre elles.
En plus des membranes stromales, les chloroplastes contiennent des thylakoïdes membranaires. Ce sont des sacs à membrane plats, fermés, en forme de disque. La taille de leur espace intermembranaire est également d'environ 20 à 30 nm. Ces thylakoïdes forment des piles comme une colonne de pièces de monnaie, appelées grana (Fig. 227). Le nombre de thylakoïdes par grana varie considérablement : de quelques-uns à 50 ou plus. La taille de ces empilements peut atteindre 0,5 micron, de sorte que les grains sont visibles sur certains objets au microscope optique. Le nombre de grains dans les chloroplastes des plantes supérieures peut atteindre 40 à 60. Les thylakoïdes du grana sont proches les uns des autres, de sorte que les couches externes de leurs membranes sont étroitement reliées ; à la jonction des membranes thylakoïdes, une couche dense d'environ 2 nm d'épaisseur se forme. En plus des chambres fermées des thylakoïdes, le grana comprend généralement également des sections de lamelles, qui forment également des couches denses de 2 nm aux points de contact de leurs membranes avec les membranes des thylakoïdes. Les lamelles stromales semblent ainsi relier les grains individuels du chloroplaste entre eux. Cependant, les cavités des chambres thylakoïdes sont toujours fermées et ne passent pas dans les chambres de l'espace intermembranaire des lamelles stromales. Les lamelles stromales et les membranes thylakoïdes sont formées par séparation de la membrane interne au cours des premiers stades du développement des plastes.
La matrice (stroma) des chloroplastes contient des molécules d'ADN et des ribosomes ; C’est également là que se produit le dépôt primaire du polysaccharide de réserve, l’amidon, sous forme de grains d’amidon.
Fonctions des chloroplastes. Les processus photosynthétiques se produisent dans les chloroplastes, conduisant à la fixation du dioxyde de carbone, à la libération d'oxygène et à la synthèse de sucres.
Un trait caractéristique des chloroplastes est la présence de pigments, les chlorophylles, qui donnent de la couleur aux plantes vertes. Grâce à la chlorophylle, les plantes vertes absorbent l’énergie du soleil et la convertissent en énergie chimique.
Le principal processus final ici est la liaison du dioxyde de carbone, l'utilisation de l'eau pour former divers glucides et la libération d'oxygène. Les molécules d'oxygène, libérées lors de la photosynthèse chez les plantes, se forment en raison de l'hydrolyse d'une molécule d'eau. Le processus de photosynthèse est une chaîne complexe composée de deux phases : claire et sombre. La première, qui se produit uniquement dans la lumière, est associée à l'absorption de la lumière par les chlorophylles et à la conduite d'une réaction photochimique (réaction de Hill). Dans la deuxième phase, qui se déroule dans l’obscurité, le CO2 est fixé et réduit, conduisant à la synthèse de glucides.
Grâce à la phase lumineuse, l'ATP est synthétisé et le NADP (nicotinamide adénine dinucléotide phosphate) est réduit, qui sont ensuite utilisés dans la réduction du CO2 dans la synthèse des glucides déjà dans la phase sombre de la photosynthèse.
Dans la phase sombre de la photosynthèse, en raison de la réduction de l’énergie du NADP et de l’ATP, le CO2 atmosphérique est lié, ce qui conduit à la formation de glucides. Ce processus de fixation du CO2 et de formation des glucides comprend de nombreuses étapes dans lesquelles un grand nombre d'enzymes sont impliquées (cycle de Calvin).
Dans le stroma des chloroplastes, les nitrites sont réduits en ammoniac grâce à l'énergie des électrons activés par la lumière ; chez les plantes, cet ammoniac sert de source d'azote lors de la synthèse des acides aminés et des nucléotides.
Ontogenèse et réarrangements fonctionnels des plastes. L'augmentation du nombre de chloroplastes et la formation d'autres formes de plastes (leucoplastes et chromoplastes) sont considérées comme un moyen de conversion des structures précurseurs, proplastide. L'ensemble du processus de développement des divers plastes semble se dérouler dans une direction, une série de changements de formes :
Proplastida ® leucoplast ® chloroplast ® chromoplaste
¯ amyloplaste¾¾¾¾¾¾¾¾¾¾
Le caractère irréversible des transitions havegénétiques des plastes a été établi. Chez les plantes supérieures, l'émergence et le développement des chloroplastes se font par le biais de modifications des proplastes (Fig. 231).
Les proplastes sont de petites vésicules à double membrane (0,4-1 μm) qui diffèrent des vacuoles cytoplasmiques par leur contenu plus dense et la présence de deux membranes délimitatrices, externe et interne (comme les promitochondries dans les cellules de levure). La membrane interne peut se plier légèrement ou former de petites vacuoles. Les proplastes se trouvent le plus souvent dans les tissus végétaux en division (cellules du méristème des racines, des feuilles, points de croissance des tiges, etc.). Une augmentation de leur nombre se produit par division ou bourgeonnement, séparation de petites vésicules à double membrane du corps du proplastide.
Le sort de ces proplastes dépend des conditions de développement des plantes. Sous un éclairage normal, les proplastes se transforment en chloroplastes. Premièrement, ils se développent, avec la formation de plis membranaires situés longitudinalement à partir de la membrane interne. Certains d'entre eux s'étendent sur toute la longueur du plaste et forment des lamelles stromales ; d'autres forment des lamelles thylakoïdes, qui sont empilées pour former le grana des chloroplastes matures.
Dans l'obscurité, les semis connaissent initialement une augmentation du volume des plastes, des étioplastes, mais le système de membranes internes ne construit pas de structures lamellaires, mais forme une masse de petites vésicules qui s'accumulent dans des zones séparées et peuvent même former des structures de réseau complexes (prolamellaires corps). Les membranes des étioplastes contiennent de la protochlorophylle, un précurseur jaune de la chlorophylle. Lorsque les cellules sont éclairées, les vésicules et les tubes membranaires se réorganisent rapidement et à partir d'eux se développe un système complet de lamelles et de thylacoïdes, caractéristiques d'un chloroplaste normal.
Les leucoplastes, contrairement aux chloroplastes, n'ont pas de système lamellaire développé (Fig. 226 b). On les retrouve dans les cellules des tissus de stockage. En raison de leur morphologie indéterminée, les leucoplastes sont difficiles à distinguer des proplastes et parfois des mitochondries. Comme les proplastes, ils sont pauvres en lamelles, mais sont néanmoins capables de former des structures thylakoïdes normales sous l'influence de la lumière et d'acquérir une couleur verte. Dans l'obscurité, les leucoplastes peuvent accumuler diverses substances de réserve dans les corps prolamellaires et des grains d'amidon secondaire se déposent dans le stroma des leucoplastes. Si l'amidon dit transitoire se dépose dans les chloroplastes, qui n'est présent ici que lors de l'assimilation du CO2, un véritable stockage de l'amidon peut alors se produire dans les leucoplastes. Dans certains tissus (endosperme des céréales, rhizomes et tubercules), l'accumulation d'amidon dans les leucoplastes conduit à la formation d'amyloplastes, entièrement remplis de granules d'amidon de réserve situés dans le stroma du plaste (Fig. 226c).
Une autre forme de plastes chez les plantes supérieures est chromoplaste, généralement coloré en jaune en raison de l'accumulation de caroténoïdes (Fig. 226d). Les chromoplastes sont formés à partir de chloroplastes et beaucoup moins fréquemment à partir de leurs leucoplastes (par exemple dans les racines de carottes). Le processus de blanchiment et de modification des chloroplastes est facilement observé lors du développement des pétales ou lors de la maturation des fruits. Dans ce cas, des gouttelettes de couleur jaune (globules) peuvent s'accumuler dans les plastes ou des corps sous forme de cristaux peuvent y apparaître. Ces processus sont provoqués par une diminution progressive du nombre de membranes dans le plaste, avec disparition de la chlorophylle et de l'amidon. Le processus de formation de globules colorés s'explique par le fait que lorsque les lamelles des chloroplastes sont détruites, des gouttelettes lipidiques sont libérées dans lesquelles divers pigments (par exemple les caroténoïdes) sont bien dissous. Ainsi, les chromoplastes sont des formes dégénératives de plastes, sujettes à la lipophanérose - la désintégration des complexes lipoprotéiques.
Structures photosynthétiques des cellules eucaryotes et procaryotes inférieures. La structure des plastes des plantes photosynthétiques inférieures (algues vertes, brunes et rouges) est en général similaire à celle des chloroplastes des cellules des plantes supérieures. Leurs systèmes membranaires contiennent également des pigments photosensibles. Les chloroplastes des algues vertes et brunes (parfois appelés chromatophores) possèdent également des membranes externe et interne ; ces derniers forment des sacs plats disposés en couches parallèles ; on ne trouve pas de granae dans ces formes (Fig. 232). Chez les algues vertes, le chromatophore comprend pyrénoïdes, représentant une zone entourée de petites vacuoles autour desquelles se dépose l'amidon (Fig. 233).
La forme des chloroplastes des algues vertes est très diversifiée - il s'agit soit de longs rubans en spirale (Spirogira), de réseaux (Oedogonium) ou de petits ronds, semblables aux chloroplastes des plantes supérieures (Fig. 234).
Parmi les organismes procaryotes, de nombreux groupes possèdent des appareils photosynthétiques et ont donc une structure particulière. Il est caractéristique des micro-organismes photosynthétiques (algues bleu-vert et de nombreuses bactéries) que leurs pigments photosensibles soient associés à la membrane plasmique ou à ses excroissances dirigées profondément dans la cellule.
En plus de la chlorophylle, les membranes des algues bleu-vert contiennent des pigments phycobilines. Les membranes photosynthétiques des algues bleu-vert forment des sacs plats (lamelles) disposés parallèlement les uns aux autres, formant parfois des piles ou des spirales. Toutes ces structures membranaires sont formées par des invaginations de la membrane plasmique.
Chez les bactéries photosynthétiques (Chromatium), les membranes forment de petites vésicules dont le nombre est si grand qu'elles remplissent la quasi-totalité du cytoplasme.
Génome des plastes. Comme les mitochondries, les chloroplastes possèdent leur propre système génétique qui assure la synthèse d’un certain nombre de protéines au sein des plastes eux-mêmes. L'ADN, divers ARN et ribosomes se trouvent dans la matrice chloroplastique. Il s'est avéré que l'ADN des chloroplastes diffère fortement de l'ADN du noyau. Il est représenté par des molécules cycliques mesurant jusqu'à 40 à 60 microns de longueur, avec un poids moléculaire de 0,8 à 1,3 x 108 daltons. Il peut y avoir plusieurs copies d’ADN dans un chloroplaste. Ainsi, dans un chloroplaste de maïs individuel, il existe 20 à 40 copies de molécules d'ADN. La durée du cycle et le taux de réplication de l’ADN nucléaire et chloroplastique, comme cela a été démontré dans les cellules d’algues vertes, ne coïncident pas. L'ADN des chloroplastes n'est pas complexé aux histones. Toutes ces caractéristiques de l'ADN chloroplastique sont proches des caractéristiques de l'ADN des cellules procaryotes. Par ailleurs, la similitude de l'ADN des chloroplastes et des bactéries est également renforcée par le fait que les principales séquences régulatrices de la transcription (promoteurs, terminateurs) sont les mêmes. Tous les types d'ARN (messager, de transfert, ribosomal) sont synthétisés sur l'ADN chloroplastique. L'ADN chloroplastique code pour l'ARNr, qui fait partie des ribosomes de ces plastes, qui appartiennent au type procaryote 70S (contiennent des ARNr 16S et 23S). Les ribosomes chloroplastiques sont sensibles à l'antibiotique chloramphénicol, qui inhibe la synthèse des protéines dans les cellules procaryotes.
La séquence entière des nucléotides dans la molécule d'ADN cyclique des chloroplastes des plantes supérieures a été complètement déchiffrée. Cet ADN peut coder jusqu'à 120 gènes, parmi lesquels : les gènes de 4 ARN ribosomiques, 20 protéines ribosomales des chloroplastes, les gènes de certaines sous-unités de l'ARN polymérase des chloroplastes, plusieurs protéines des photosystèmes I et II, 9 des 12 sous-unités de l'ATP synthétase, des parties de protéines des complexes de la chaîne de transport d'électrons, l'une des sous-unités de la ribulose diphosphate carboxylase (l'enzyme clé pour la liaison du CO2), 30 molécules d'ARNt et 40 autres protéines encore inconnues. Il est intéressant de noter qu'un ensemble similaire de gènes dans l'ADN des chloroplastes a été trouvé chez des représentants aussi éloignés de plantes supérieures que le tabac et la mousse de foie.
La majeure partie des protéines chloroplastiques est contrôlée par le génome nucléaire. Un certain nombre de protéines et d'enzymes les plus importantes et, par conséquent, les processus métaboliques des chloroplastes sont sous le contrôle génétique du noyau. La plupart des protéines ribosomales sont sous le contrôle de gènes nucléaires. Toutes ces données parlent des chloroplastes comme de structures à autonomie limitée.
4.6. Cytoplasme : système musculo-squelettique (cytosquelette)
Toutes les nombreuses réactions motrices d’une cellule reposent sur des mécanismes moléculaires communs. De plus, la présence d'appareils moteurs est combinée et structurellement liée à l'existence de formations intracellulaires de support, de charpente ou squelettiques. Par conséquent, ils parlent du système cellulaire musculo-squelettique.
Les composants du cytosquelette comprennent des complexes ou filaments protéiques filamenteux et non ramifiés (filaments minces).
Il existe trois groupes de filaments, différant à la fois par leur composition chimique et leur ultrastructure, ainsi que par leurs propriétés fonctionnelles. Les fils les plus fins sont des microfilaments ; leur diamètre est d'environ 8 nm et ils sont principalement constitués de protéine actine. Un autre groupe de structures filamenteuses sont les microtubules, qui ont un diamètre de 25 nm et sont constitués principalement de la protéine tubuline, et enfin des filaments intermédiaires d'un diamètre d'environ 10 nm (intermédiaire par rapport à 6 nm et 25 nm), formés de différents mais liés. protéines (Fig. 238, 239).
Toutes ces structures fibrillaires sont impliquées dans les processus de mouvement physique des composants cellulaires ou même des cellules entières, dans certains cas elles jouent un rôle purement squelettique. Les éléments cytosquelettiques se retrouvent dans toutes les cellules eucaryotes sans exception ; des analogues de ces structures fibrillaires se trouvent également chez les procaryotes.
Les propriétés générales des éléments du cytosquelette sont qu'ils sont des polymères fibrillaires protéiques non ramifiés, instables, capables de polymérisation et de dépolymérisation, qui conduisent à la motilité cellulaire, par exemple à des modifications de la forme des cellules. Les composants du cytosquelette, avec la participation de protéines supplémentaires spéciales, peuvent stabiliser ou former des assemblages fibrillaires complexes et jouer uniquement un rôle d'échafaudage. Lorsqu'elles interagissent avec d'autres protéines translocatrices spéciales (ou protéines motrices), elles participent à divers mouvements cellulaires.
Selon leurs propriétés et fonctions, les éléments du cytosquelette sont divisés en deux groupes : uniquement les fibrilles du cadre - les filaments intermédiaires, et les fibrilles musculo-squelettiques - les microfilaments d'actine interagissant avec les protéines motrices - les myosines, et les microtubules de tubuline interagissant avec les protéines motrices, les dynéines et les kinésines.
Le deuxième groupe de fibrilles cytosquelettiques (microfilaments et microtubules) propose deux modes de mouvement fondamentalement différents. Le premier d’entre eux repose sur la capacité de la principale protéine des microfilaments, l’actine, et de la principale protéine des microtubules, la tubuline, à polymériser et dépolymériser. Lorsque ces protéines se lient à la membrane plasmique, ses modifications morphologiques s'observent sous la forme de formation d'excroissances (pseudopodes et lamellipodes) au bord de la cellule.
Dans une autre méthode de mouvement, les fibrilles d'actine (microfilaments) ou de tubuline (microtubules) sont des structures directrices le long desquelles se déplacent des protéines mobiles spéciales - les moteurs. Ces dernières peuvent se lier aux composants membranaires ou fibrillaires de la cellule et ainsi participer à leur mouvement.
Ce sont des corps incolores ou colorés dans le protoplasme des cellules végétales, représentant un système complexe de membranes internes (organites membranaires) et remplissant diverses fonctions. Les plastes incolores sont appelés leucoplastes, les plastes de couleurs différentes (jaune, orange ou rouge) sont appelés chromoplastes et les verts sont appelés chloroplastes. La cellule des plantes supérieures contient environ 40 chloroplastes dans lesquels se produit la photosynthèse. Comme déjà mentionné, ils sont capables de se reproduire de manière autonome, indépendamment de la division cellulaire. La taille et la forme des mitochondries et des chloroplastes, la présence d'ADN circulaire double brin et leurs propres ribosomes dans leur matrice rendent ces organites similaires aux cellules bactériennes. Il existe une théorie sur l'origine symbiotique de la cellule eucaryote, selon laquelle les ancêtres des mitochondries et des chloroplastes modernes étaient autrefois des organismes procaryotes indépendants.
Les plastes ne sont caractéristiques que des plantes. On ne les trouve pas chez les champignons ni chez la plupart des animaux, à l'exception de certains protozoaires photosynthétiques.
Les précurseurs des plastes sont les proplastes, petites formations généralement incolores trouvées dans les cellules en division des racines et des pousses. Si le développement des proplastes en structures plus différenciées est retardé en raison du manque de lumière, un ou plusieurs corps prolamellaires (amas de membranes tubulaires) peuvent y apparaître. Ces plastes incolores sont appelés étioplastes. Les étioplastes se transforment en chloroplastes à la lumière et les thylakoïdes se forment à partir des membranes des corps prolamellaires. Selon la couleur associée à la présence ou à l'absence de certains pigments, il existe trois principaux types de plastes (voir ci-dessus) : les chloroplastes, les chromoplastes et les leucoplastes. Généralement, un seul type de plaste se trouve dans une cellule. Cependant, il a été établi que certains types de plastes peuvent se transformer en d’autres.
Les plastes sont des formations cellulaires relativement grandes. Les plus gros d'entre eux - les chloroplastes - atteignent 4 à 10 microns de longueur chez les plantes supérieures et sont clairement visibles au microscope optique. La forme des plastes colorés est le plus souvent lenticulaire ou elliptique. En règle générale, on trouve plusieurs dizaines de plastes dans les cellules, mais chez les algues, où les plastes sont souvent grands et de formes variées, leur nombre est parfois faible (1 à 5). Ces plastes sont appelés chromatophores. Les leucoplastes et les chromoplastes peuvent avoir des formes différentes.
La fonction principale des chloroplastes est la photosynthèse. Le rôle central dans ce processus appartient à la chlorophylle, ou plus précisément à plusieurs de ses modifications. Les réactions lumineuses de la photosynthèse s'effectuent principalement dans le grana, les réactions sombres - dans le stroma
- (du grec plastos façonné) organites cytoplasmiques des cellules végétales. Ils contiennent souvent des pigments qui déterminent la couleur du plaste. Les plantes supérieures ont des plastes verts, des chloroplastes, des leucoplastes incolores et des chromoplastes de couleurs variées ; ... Grand dictionnaire encyclopédique
PLASTIDES- (du grec plastides créant, formant, à partir de plastos sculptés, façonnés), des organites eucaryotes, des cellules. Bien visible au microscope optique. Chaque P. est limité par deux membranes élémentaires ; pour beaucoup, c'est typique b. ou m... ... Dictionnaire encyclopédique biologique
PLASTIDES- PLASTIDES, BIOPLASTES ou LEUCITES Composant morphologique des cellules végétales, constitué. d'un nombre important de corps de différentes tailles et formes, couchés. près du noyau. Dictionnaire de mots étrangers inclus dans la langue russe. Chudinov A.N. ... Dictionnaire des mots étrangers de la langue russe
Plastides- *plaste* Les plastes sont des organites spécifiques auto-réplicants (voir), localisés dans le cytoplasme des cellules végétales eucaryotes. En fonction de leur capacité à lier les pigments et de leurs caractéristiques fonctionnelles, les P. sont divisés en incolores... ... La génétique. Dictionnaire encyclopédique
plastes- (du grec plastós façonné), organites cytoplasmiques des cellules végétales. Ils contiennent souvent des pigments qui provoquent la couleur des plastes. Les plantes supérieures ont des plastes verts, des chloroplastes, des leucoplastes incolores, de couleurs variées... ... Dictionnaire encyclopédique
Plastides- (du grec plástides créant, formant, à partir de plastós façonnés, façonnés) des organites intracellulaires du cytoplasme des plantes autotrophes, contenant des pigments et réalisant la synthèse de substances organiques. Dans les plantes supérieures, il existe 3 types de P... Grande Encyclopédie Soviétique
plastes- Les statuts des plastides T sritis augalininkystė apibrėžtis Bespalviai arba spalvoti organoidai, esantys autotrofinių augalų citoplazmoje ir atliekantys organinių medžiagų (krakmolo, riebalų ar baltymų) sintezę. Pagal pigmentacijos ir funkcijos… … Nous vous recommandons d'augmenter les sélections dans les kystes sékliniques des extrémités žodynas
Plastides- sinon les leucites sont un composant morphologique des cellules végétales. Outre le plasma et le noyau, ces derniers (à l'exception des champignons) contiennent généralement un nombre plus ou moins important de corps de tailles et de formes diverses, situés dans... ... Dictionnaire encyclopédique F.A. Brockhaus et I.A. Éfron
PLASTIDES- (du grec plastos façonné), cytoplasmique. fait pousser des organites. cellules. Contiennent souvent des pigments qui déterminent la couleur de P. En supérieur. r nii vert P. chloroplastes, leucoplastes incolores, chromoplastes de couleurs variées ; dans la plupart des algues... Sciences naturelles. Dictionnaire encyclopédique
plastes- des organites cellulaires à double membrane incolores ou colorés qui possèdent leur propre ADN et ribosomes, ainsi qu'un système thylakoïde exprimé à des degrés divers. Capable de se reproduire en divisant en deux. Effectuer différentes fonctions. Dans les cellules supérieures... ... Anatomie et morphologie des plantes
Depuis l'école. Le cours de botanique indique que dans les cellules végétales, les plastes peuvent avoir différentes formes, tailles et remplir différentes fonctions dans la cellule. Cet article rappellera à ceux qui ont obtenu leur diplôme il y a longtemps la structure des plastes, leurs types et leurs fonctions, et sera utile à tous ceux qui s'intéressent à la biologie.
Structure
L'image ci-dessous montre schématiquement la structure des plastes dans une cellule. Quel que soit son type, il possède une membrane externe et interne qui remplit une fonction protectrice, le stroma est un analogue du cytoplasme, des ribosomes, d'une molécule d'ADN et des enzymes.
Les chloroplastes contiennent des structures spéciales - grana. Les grana sont formés de thylakoïdes, des structures en forme de disque. Les thylakoïdes participent et oxygènent.
Dans les chloroplastes, les grains d'amidon se forment à la suite de la photosynthèse.
Les leucoplastes ne sont pas pigmentés. Ils ne contiennent pas de thylakoïdes et ne participent pas à la photosynthèse. La plupart des leucoplastes sont concentrés dans la tige et la racine de la plante.
Les chromoplastes contiennent des gouttelettes lipidiques - des structures contenant des lipides nécessaires pour fournir de l'énergie supplémentaire à la structure plastidienne.
Les plastes peuvent être de différentes couleurs, tailles et formes. Leurs tailles varient de 5 à 10 microns. La forme est généralement ovale ou ronde, mais peut être toute autre.
Types de plastes
Les plastes peuvent être incolores (leucoplastes), verts (chloroplastes), jaunes ou oranges (chromoplastes). Ce sont les chloroplastes qui donnent aux feuilles des plantes leur couleur verte.

Une autre variété est responsable de la coloration jaune, rouge ou orange.
Les plastes incolores de la cellule servent de stockage des nutriments. Les leucoplastes contiennent des graisses, de l'amidon, des protéines et des enzymes. Lorsque la plante a besoin d'énergie supplémentaire, l'amidon est décomposé en monomères : le glucose.
Les leucoplastes, dans certaines conditions (sous l'influence de la lumière du soleil ou lors de l'ajout de produits chimiques), peuvent se transformer en chloroplastes, les chloroplastes se transforment en chromoplastes lorsque la chlorophylle est détruite et les pigments colorants des chromoplastes - carotène, anthocyanine ou xanthophylle - commencent à prédominer en couleur. Cette transformation est perceptible en automne, lorsque les feuilles et de nombreux fruits changent de couleur en raison de la destruction de la chlorophylle et de l'apparition de pigments chromoplastiques.

Les fonctions
Comme indiqué ci-dessus, les plastes se présentent sous de nombreuses formes différentes et leurs fonctions dans une cellule végétale dépendent de la variété.
Les leucoplastes servent principalement à stocker les nutriments et à maintenir la vie de la plante grâce à leur capacité à stocker et à synthétiser des protéines, des lipides et des enzymes.
Les chloroplastes jouent un rôle clé dans le processus de photosynthèse. Avec la participation du pigment chlorophyllien concentré dans les plastes, les molécules de dioxyde de carbone et d'eau sont converties en molécules de glucose et d'oxygène.
Les chromoplastes, en raison de leur couleur vive, attirent les insectes pour polliniser les plantes. Les recherches sur les fonctions de ces plastes sont toujours en cours.
Nous recommandons également
 Fracture : ce que vous pouvez et ne pouvez pas faire
Fracture : ce que vous pouvez et ne pouvez pas faire
 Infinitif en anglais
Infinitif en anglais
 Caractéristiques de la fraise de "L'Inspecteur général" N
Caractéristiques de la fraise de "L'Inspecteur général" N
 Qu'est-ce qu'un plaste en biologie ?
Qu'est-ce qu'un plaste en biologie ?
 Relevé bancaire du compte courant : qu'est-ce que c'est et ses caractéristiques
Relevé bancaire du compte courant : qu'est-ce que c'est et ses caractéristiques
 Liste des instructions de base pour la comptabilité budgétaire
Liste des instructions de base pour la comptabilité budgétaire