Kako so snovi vključene v glukoneogenezo? Sinteza glukoze iz aminokislin Glukoneogeneza iz glutaminske kisline
Iz krvi v tkivne celice vstopi glukoza po mehanizmu olajšane difuzije s sodelovanjem nosilnih beljakovin. Izjema so celice mišic in maščobnega tkiva, kjer olajšano difuzijo uravnava inzulin (hormon trebušne slinavke). V odsotnosti inzulina je plazemska membrana teh celic neprepustna za glukozo, saj ne vsebuje beljakovin nosilca glukoze.
Prenosniki glukoze(GLUT) najdemo v vseh tkivih.
|
Vrste GLUT |
Lokalizacija v organih |
|
Predvsem v možganih, posteljici, ledvicah, debelem črevesu |
|
|
Predvsem v jetrih, ledvicah, β-celicah Langerhansovih otočkov, enterocitih |
|
|
V mnogih tkivih, vključno z možgani, posteljico, ledvicami |
|
|
V mišicah (skeletnih, srčnih), maščobnem tkivu |
|
|
(odvisno od insulina) |
V odsotnosti insulina skoraj v celoti v citoplazmi |
|
V tankem črevesu. Morda nosilec fruktoze. |
Opisanih 5 vrst GLUT ima podobno primarno strukturo in organizacijo domene.
GLUT-1 zagotavlja stalen pretok glukoze v možgane;
GLUT-2 najdemo v celicah organov, ki sproščajo glukozo v kri. S sodelovanjem GLUT-2 se glukoza prenaša v kri iz enterocitov in jeter. GLUT-2 sodeluje pri transportu glukoze do β-celic trebušne slinavke;
GLUT-3 ima večjo afiniteto za glukozo kot GLUT-1. Prav tako zagotavlja stalen pretok glukoze v celice živčnega in drugih tkiv;
GLUT-4 je glavni prenašalec glukoze v celice mišic in maščobnega tkiva;
GLUT-5 se nahaja predvsem v celicah tankega črevesa. Njegove funkcije niso dobro znane.
Vse vrste GLUT najdemo tako v plazemski membrani kot v membranskih veziklih v citoplazmi. Vendar je le GLUT-4, lokaliziran v citoplazmatskih veziklih, vgrajen v plazemsko membrano celic mišičnega in maščobnega tkiva s sodelovanjem hormona trebušne slinavke inzulina. Ker je oskrba mišic in maščobnega tkiva z glukozo odvisna od insulina, se ta tkiva imenujejo insulinsko odvisna.
Učinek inzulina na gibanje prenašalcev glukoze iz citoplazme v plazemsko membrano.
1 - vezava insulina na receptor; 2 - odsek insulinskega receptorja, obrnjen proti notranjosti celice, spodbuja gibanje transporterjev glukoze; 3, 4 - transporterji v sestavi veziklov, ki jih vsebujejo, se premikajo na plazemsko membrano celice, so vključeni v njeno sestavo in prenašajo glukozo v celico.
Poznane so različne nepravilnosti pri delu prenašalcev glukoze. Podedovana okvara teh beljakovin je lahko osnova za diabetes mellitus, ki ni odvisen od insulina. Motnje delovanja GLUT-4 so možne v naslednjih fazah:
prenos insulinskega signala o premiku tega transporterja na membrano;
gibanje transporterja v citoplazmi;
vključitev v sestavo membrane;
ločitev od membrane itd.
Glukoneogeneza je sinteza glukoze iz neogljikovih hidratov: laktat, piruvat, glicerol, ketonske kisline Krebsovega cikla in drugih ketonskih kislin, od amino kisline... Vse aminokisline, razen ketogenega levcina in lizina, lahko sodelujejo pri sintezi glukoze. Ogljikovi atomi nekaterih od njih (glukogenih) so v celoti vgrajeni v molekulo glukoze, nekateri (pomešani) delno.
Glukoneogeneza poleg pridobivanja glukoze zagotavlja tudi čiščenje laktat nenehno nastaja v rdečih krvnih celicah ali med delom mišic, in glicerol, ki je produkt lipolize v maščobnem tkivu.
Kot veste, v glikoliza obstajajo tri nepopravljive reakcije: piruvat kinaza(deseti), fosfofruktokinaza(tretji) in heksokinaza(prvi). Pri teh reakcijah se sprosti energija za sintezo ATP. Zato v obratnem procesu energijske ovire, ki ga celica obide s pomočjo dodatnih reakcij.
Glukoneogeneza vključuje vse reverzibilne reakcije glikoliza in posebne rešitve, tj. ne ponovi popolnoma reakcije oksidacije glukoze. Njegove reakcije se lahko odvijajo v vseh tkivih, razen zadnje reakcije glukoza-6-fosfataze, ki se pojavi le v jetra in ledvice... Zato se, strogo gledano, glukoneogeneza pojavlja le v teh dveh organih.
Na tej stopnji glukoneogeneze delujeta dva ključna encima – v mitohondrijih piruvat karboksilaze in v citosolu fosfoenolpiruvat karboksikinaza,v tem primeru reakcije porabijo dva makroerga - ATP in GTP.
Kemično je rešitev za deseto reakcijo precej preprosta:
Poenostavljena različica obvoda desete reakcije glikolize
Vendar je bistvo v tem piruvat karboksilaze se nahaja v mitohondriju in - v citosolu. Dopolnjuje problem neprepustnost mitohondrijska membrana za oksaloacetat... Toda skozi membrano lahko prehaja malat, predhodnik oksaloacetata po CTA.
Zato je v resnici vse videti bolj zapleteno:
1. V citosolu se lahko med oksidacijo pojavi pirovična kislina mlečna kislina in v reakciji transaminacije alanin... Po tem piruvat z ioni H +, ki se premikajo vzdolž protonskega gradienta, prodre v mitohondrije. V mitohondrijih piruvat karboksilaze pretvori pirovično kislino v oksaloacetat.
Reakcija piruvat karboksilaze se nenehno pojavlja v celici, saj je oksaloacetat glavni regulator hitrosti CTX. Reakcija se imenuje anaplerotični (obnovitveni) odziv CTX.
2. Nadalje se oksaloacetat lahko pretvori v fosfoenolpiruvat, vendar mora za to najprej vstopiti v citosol. Zato se reakcija redukcije oksaloacetata v malat pojavi s sodelovanjem malat dehidrogenaza... Posledično se malat kopiči, gre v citosol in se tu pretvori nazaj v oksaloacetat.
Presežek NADH v mitohondrijih omogoča obrnitev malat dehidrogenazne reakcije CTX. NADH izvira iz β-oksidacije maščobnih kislin, ki se med postom aktivira v hepatocitih.
3. V citoplazmi fosfoenolpiruvat karboksikinaza izvaja transformacijo oksaloacetata v fosfoenolpiruvat, reakcija zahteva energijo GTP. Isti ogljik se odcepi od molekule, ko je pritrjen.

Obhod desete reakcije glikolize
Mimo tretje reakcije glikolize
Drugo oviro za sintezo glukoze - fosfofruktokinazno reakcijo - premagamo s pomočjo encima. fruktoza-1,6-difosfataza... Ta encim se nahaja v ledvicah, jetrih, progastih mišicah. Tako so ta tkiva sposobna sintetizirati fruktozo-6-fosfat in glukozo-6-fosfat.
Glukoneogeneza je proces sinteze glukoze iz neogljikovih hidratov. Pri sesalcih to funkcijo opravljajo predvsem jetra , v manjši meri - ledvice in celice črevesne sluznice ... Glavni substrati glukoneogeneze so piruvat, laktat, glicerin, aminokisline (slika 10).
Slika 10
Glukoneogeneza zagotavlja potrebe telesa po glukozi v primerih, ko prehrana vsebuje premajhno količino ogljikovih hidratov (telesna aktivnost, post). Stalna oskrba z glukozo je še posebej potrebna za živčni sistem in rdeče krvne celice. Ko koncentracija glukoze v krvi pade pod določeno kritično raven, je delovanje možganov moteno; pri hudi hipoglikemiji nastopi koma, ki je lahko usodna.
Telo ima dovolj zalog glikogena, da zadovolji potrebe po glukozi med obroki. Z ogljikovimi hidrati ali popolnim stradanjem, pa tudi v pogojih dolgotrajnega fizičnega dela, se koncentracija glukoze v krvi vzdržuje zaradi glukoneogeneze. Ta proces lahko vključuje snovi, ki se lahko pretvorijo v piruvat ali kateri koli drug metabolit glukoneogeneze. Slika prikazuje točke vključitve primarnih substratov v glukoneogenezo:
Glukoza je nujna za maščobno tkivo kot vir glicerola, ki je del gliceridov; igra bistveno vlogo pri ohranjanju učinkovitih koncentracij metabolitov cikla citronske kisline v številnih tkivih. Tudi v razmerah, ko večino potreb telesa po kalorijah pokriva maščoba, vedno obstaja določena potreba po glukozi. Poleg tega je glukoza edino gorivo za delovanje skeletnih mišic v anaerobnih pogojih. Je predhodnik mlečnega sladkorja (laktoze) v mlečnih žlezah in ga plod med razvojem aktivno porablja. Mehanizem glukoneogeneze se uporablja za odstranjevanje produktov presnove tkiva iz krvi, na primer laktata, ki nastaja v mišicah in eritrocitih, glicerola, ki se nenehno tvori v maščobnem tkivu.
Vključitev različnih substratov v glukoneogenezo je odvisna od fiziološkega stanja organizma. laktat je produkt anaerobne glikolize v eritrocitih in delujočih mišicah. glicerol sprošča med hidrolizo maščob v maščobnem tkivu v postabsorpcijskem obdobju ali med fizičnim naporom. Amino kisline nastanejo kot posledica razgradnje mišičnih beljakovin.
Sedem reakcij glikolize je zlahka reverzibilnih in se uporabljajo pri glukoneogenezi. Toda tri kinazne reakcije so ireverzibilne in jih je treba ustaviti (slika 12). Tako sta fruktoza-1,6-difosfat in glukoza-6-fosfat defosforilirana s posebnimi fosfatazami, piruvat pa se fosforilira, da tvori fosfoenolpiruvat v dveh vmesnih korakih prek oksaloacetata. Nastajanje oksaloacetata katalizira piruvat karboksilaza. Ta encim vsebuje biotin kot koencim. Oksaloacetat nastaja v mitohondrijih, se prenaša v citosol in sodeluje pri glukoneogenezi. Opozoriti je treba, da vsaka od ireverzibilnih reakcij glikolize skupaj z ustrezno ireverzibilno reakcijo glukoneogeneze tvori cikel, imenovan substrat:
Slika 12
Obstajajo trije takšni cikli - ki ustrezajo trem ireverzibilnim reakcijam. Ti cikli služijo točke uporabe regulativnih mehanizmov , zaradi česar se tok metabolitov spreminja bodisi po poti razgradnje glukoze bodisi po poti njene sinteze.
Smer reakcij prvi cikel substrata uravnava predvsem koncentracijo glukoze. Med prebavo se koncentracija glukoze v krvi dvigne. Aktivnost glukokinaze je v teh pogojih največja. Posledično se glikolitična reakcija pospeši. glukoza ® glukoza-6-fosfat. Poleg tega insulin inducira sintezo glukokinaze in s tem pospešuje fosforilacijo glukoze. Ker glukoza-6-fosfat ne zavira jetrne glukokinaze (za razliko od mišične heksokinaze), je glavnina glukoza-6-fosfata usmerjena vzdolž glikolitične poti.
Pretvorbo glukoza-6-fosfata v glukozo katalizira druga specifična fosfataza - glukoza-6-fosfataza. Prisoten je v jetrih in ledvicah, v mišičnem in maščobnem tkivu pa ga ni. Prisotnost tega encima omogoča tkivu, da dovaja glukozo v kri.
Razgradnjo glikogena s tvorbo glukoza-1-fosfata izvaja fosforilaza. Sinteza glikogena poteka po povsem drugačni poti, skozi tvorbo uridin difosfat glukoze, in je katalizirana. glikogen sintaza.
Drugi substrat cikel: pretvorba fruktozo-1,6-bisfosfata v fruktozo-6-fosfat, katalizirana s posebnim encimom fruktoza-1,6-bisfosfataza. Ta encim se nahaja v jetrih in ledvicah, najdemo pa ga tudi v progastih mišicah.
Smer reakcij drugi substrat cikel je odvisen od aktivnosti fosfofruktokinaze in fosfataze fruktozo-1,6-bisfosfata. Aktivnost teh encimov je odvisna od koncentracije fruktozo-2,6-bisfosfata.
Fruktoza-2,6-bisfosfat nastane s fosforilacijo fruktozo-6-fosfata s sodelovanjem bifunkcionalnega encima (BIF), ki tudi katalizira obratno reakcijo.
Aktivnost kinaze se pojavi, ko je bifunkcionalni encim v defosforilirani obliki (BIF-OH). Defosforilirana oblika BIF je značilna za obdobje absorpcije, ko je indeks insulin-glukagon visok.
Pri nizkem inzulinsko-glukagonskem indeksu, značilnem za obdobje daljšega posta, pride do fosforilacije BIF in manifestacije njegove fosfatazne aktivnosti, kar povzroči zmanjšanje količine fruktozo-2,6-bisfosfata, upočasnitev glikolize in prehod na glukoneogeneza.
Reakcije kinaze in fosfataze katalizirajo različni aktivni centri BIF, vendar je v vsakem od dveh stanj encima – fosforiliranega in defosforiliranega – eno od aktivnih centrov zavirano.
16.2.1. Glukoneogeneza je biosinteza glukoze iz različnih neogljikovih hidratov. Biološka vloga glukoneogeneze je vzdrževanje stalne ravni glukoze v krvi, ki je potrebna za normalno oskrbo z energijo tkiv, za katere je značilna stalna potreba po ogljikovih hidratih. To še posebej velja za centralni živčni sistem.
Vloga glukoneogeneze se poveča z nezadostnim vnosom ogljikovih hidratov iz hrane. Torej, v telesu stradajoče osebe se lahko sintetizira do 200 g glukoze na dan. Glukoneogeneza se hitreje kot drugi metabolni procesi odziva na spremembe v prehrani: vnos velikih količin beljakovin in maščob s hrano aktivira procese glukoneogeneze; presežek ogljikovih hidratov, nasprotno, zavira tvorbo glukoze.
Intenzivno telesno aktivnost spremlja hitro izčrpavanje zalog glukoze v telesu. V tem primeru je glukoneogeneza glavni način obnavljanja virov ogljikovih hidratov, ki preprečuje razvoj hipoglikemije. Glukoneogeneza v telesu je tesno povezana tudi s procesi nevtralizacije amoniaka in vzdrževanja kislinsko-bazičnega ravnovesja.
16.2.2. Glavno mesto biosinteze glukoze de novo je jetra. Glukoneogeneza se pojavi tudi v ledvični skorji. Na splošno velja, da je prispevek ledvic k glukoneogenezi v fizioloških pogojih približno 10 % glukoze, sintetizirane v telesu; pri patoloških stanjih se lahko ta delež znatno poveča. V sluznici tankega črevesa je bila ugotovljena nepomembna aktivnost encimov glukoneogeneze.
16.2.3. Zaporedje reakcij glukoneogeneze je preobrat ustreznih reakcij glikolize. Samo tri reakcije glikolize so nepopravljive zaradi znatnih energijskih premikov, ki se pojavijo med njihovim potekom:
a) fosforilacija glukoze; b) fosforilacija fruktozo-6-fosfata; c) pretvorba fosfoenolpiruvata v piruvat.Obhod teh energijskih ovir zagotavljajo ključni encimi glukoneogeneze.
Povratna pretvorba piruvata v fosfoenolpiruvat zahteva sodelovanje dveh encimov. Prvi je piruvat karboksilaze - katalizira reakcijo tvorbe oksaloacetata (slika 16.4, reakcija 1). Koencim piruvat karboksilaze je biotin (vitamin H). Reakcija teče v mitohondrijih. Njegova vloga je tudi napolniti oksaloacetatni sklad za Krebsov cikel.
Vse nadaljnje reakcije glukoneogeneze potekajo v citoplazma ... Mitohondrijska membrana je neprepustna za oksaloacetat in se v citoplazmo prenaša v obliki drugih metabolitov: malata ali aspartata. V citoplazmi se te spojine ponovno pretvorijo v oksaloacetat. V glavni vlogi fosfoenolpiruvat karboksikinaza iz oksaloacetata nastane fosfoenolpiruvat (slika 16.4, reakcija 2).
Fosfoenolpiruvat se zaradi inverzije številnih reakcij glikolize pretvori v fruktozo-1,6-difosfat. Pretvorbo fruktozo-1,6-difosfata v fruktozo-6-fosfat katalizira fruktoza difosfataza (Slika 16.4, reakcija 3).
Fruktoza-6-fosfat se izomerizira v glukozo-6-fosfat. Končna reakcija glukoneogeneze je hidroliza glukoza-6-fosfata s sodelovanjem encima glukoza-6-fosfataze (slika 16.4, reakcija 4).
Slika 16.4. Bypass reakcije glukoneogeneze .
16.2.4. Glavni viri glukoze v glukoneogenezi so laktat, aminokisline, glicerol in presnovki Krebsovega cikla.
laktat- končni produkt anaerobne oksidacije glukoze. Lahko sodeluje pri glukoneogenezi po oksidaciji v piruvat v reakciji laktat dehidrogenaze (glejte razdelek »Glikoliza«, slika 15.4, reakcija 11). Pri dolgotrajnem fizičnem delu so glavni vir laktata skeletne mišice, v celicah katerih prevladujejo anaerobni procesi. Kopičenje mlečne kisline v mišicah omejuje njihovo delovanje. To je posledica dejstva, da se s povečanjem koncentracije mlečne kisline v tkivu raven pH zmanjša (laktacidoza). Sprememba pH vodi do zaviranja encimov najpomembnejših presnovnih poti. Pri odstranjevanju nastale mlečne kisline pripada pomembno mesto Corey cikel glukoze in laktata (Slika 16.5).

Slika 16.5. Coreyev cikel in glukozno-alaninski cikel (za pojasnila glejte besedilo).
Glukogene aminokisline, ki vključuje večino beljakovinskih aminokislin. Vodilno mesto v glukoneogenezi med aminokislinami pripada Alanin , ki se lahko s transaminacijo pretvori v piruvat. Med postom, fizičnim delom in drugimi stanji v telesu deluje cikel glukoza-alanin podobno kot Coreyev cikel za laktat (slika 16.2). Obstoj cikla alanin-glukoza preprečuje zastrupitev telesa, saj v mišicah ni encimov, ki izkoriščajo amoniak. Zaradi treninga se moč tega cikla znatno poveča.
Druge aminokisline se lahko, kot je alanin, pretvorijo v piruvat in tudi v vmesni produkti Krebsovega cikla (α-ketoglutarat, fumarat, sukcinil-CoA). Vsi ti presnovki se lahko pretvorijo v oksaloacetat in sodelujejo pri glukoneogenezi.
glicerol- produkt hidrolize lipidov v maščobnem tkivu. Ta proces močno pospeši post. V jetrih se glicerol pretvori v dioksiaceton fosfat, vmesni produkt glikolize, in se lahko uporablja pri glukoneogenezi.
maščobna kislina in acetil-CoA niso predhodniki glukoze. Oksidacija teh spojin zagotavlja energijo za sintezo glukoze.
16.2.5. Energetska bilanca. Pot za sintezo glukoze iz piruvata (slika 16.6) vsebuje tri reakcije, ki jih spremlja poraba energije ATP ali GTP:
a) tvorba oksaloacetata iz piruvata (molekula ATP se porabi); b) tvorba fosfoenolpiruvata iz oksaloacetata (porabljena je molekula GTP); c) obrat fosforilacije prvega substrata - tvorba 1,3-difosfoglicerata iz 3-fosfoglicerata (molekula ATP se porabi).Vsaka od teh reakcij se ponovi dvakrat, saj se 2 molekuli piruvata (C3) uporabita za tvorbo 1 molekule glukoze (C6). Zato je energijska bilanca sinteze glukoze iz piruvata - 6 molekul nukleozid trifosfatov (4 molekule ATP in 2 molekuli GTP). Ko se uporabljajo drugi predhodniki, je energijska bilanca biosinteze glukoze drugačna.

Slika 16.6. Energetska bilanca biosinteze glukoze iz laktata.
16.2.6. Regulacija glukoneogeneze. Hitrost glukoneogeneze je določena z razpoložljivostjo substratov prekurzorjev glukoze. Povečanje koncentracije katerega koli od predhodnikov glukoze v krvi povzroči stimulacijo glukoneogeneze.
Nekateri metaboliti so alosterični efektorji encimov glukoneogeneze. Na primer, acetil-CoA pri povišanih koncentracijah alosterično aktivira piruvat karboksilazo, ki katalizira prvo reakcijo glukoneogeneze. Adenozin monofosfat, nasprotno, ima zaviralni učinek na fruktozo difosfatazo, presežek glukoze pa zavira glukozo-6-fosfatazo.
Hormon trebušne slinavke glukagon, hormona nadledvične žleze adrenalin in kortizol povečajo hitrost biosinteze glukoze v telesu, povečajo aktivnost ključnih encimov glukoneogeneze ali povečajo koncentracijo teh encimov v celicah. Hormon trebušne slinavke inzulin pomaga zmanjšati hitrost glukoneogeneze v telesu.
V piruvatu ali enem od vmesnih produktov cikla trikarboksilne kisline.
Pri vretenčarjih se glukoneogeneza najbolj intenzivno pojavlja v celicah jeter in ledvic (v skorji).
Prva faza sinteze poteka v mitohondrijih (slika 10.6). Piruvat karboksilaza, ki katalizira to reakcijo, je alosterični mitohondrijski encim. Acetil-CoA je potreben kot alosterični aktivator tega encima. Mitohondrijska membrana je neprepustna za nastali oksaloacetat. Slednji se tukaj, v mitohondrijih, reducira na malat:


Nadaljnja pretvorba oksaloacetata v fosfoenolpiruvat poteka v citosolu celice.
Pretvorba fruktozo-1,6-bisfosfata v fruktozo-6-fosfat... Fosfo-enolpiruvat, ki nastane iz piruvata, se pretvori v fruktozo-1,6-bisfosfat kot rezultat vrste reverzibilnih reakcij glikolize. Temu sledi reakcija fosfofruktokinaze, ki je nepovratna. Glukoneogeneza zaobide to endergonično reakcijo. Pretvorbo fruktozo-1,6-bis-fosfata v fruktozo-6-fosfat katalizira specifična fosfataza:

riž. 10.6. Tvorba fosfoenol piruvata iz piruvata. 1 - piruvat karboksilaza; 2 - malat dehidrogenaza (mitohondrijska); 3-malat dehidrogenaza (citoplazmatska); 4 - fosfoenolpiruvat karboksi kinaza.

Druga pomembna točka pri regulaciji glukoneogeneze je reakcija, ki jo katalizira fruktoza-1,6-bisfosfataza, encim, ki ga zavira AMP. AMP ima nasproten učinek na fosfofruktokinazo, torej je za ta encim alosterični aktivator. Z nizko koncentracijo AMP in visoko stopnjo ATP se stimulira glukoneogeneza. Nasprotno, ko je vrednost razmerja ATP / AMP majhna, se v celici opazi razgradnja glukoze.
Leta 1980 je skupina belgijskih raziskovalcev (G. Hers in drugi) v jetrnem tkivu odkrila fruktozo-2,6-bisfosfat, ki je močan regulator aktivnosti dveh naštetih encimov:

Fruktoza-2,6-bisfosfat aktivira fosfofruktokinazo in zavira fruktozo-1,6-bisfosfatazo. Povečanje ravni fruktoze-2,6-bis-fosfata v celici spodbuja povečano glikolizo in zmanjšanje hitrosti glukoneogeneze. Z zmanjšanjem koncentracije fruktozo-2,6-bisfosfata opazimo nasprotno sliko.

Pokazalo se je tudi, da je bifunkcionalni encim po drugi strani reguliran s fosforilacijo, odvisno od cAMP. Fosforilacija vodi do povečanja aktivnosti fosfataze in zmanjšanja aktivnosti fosfokinaze bifunkcionalnega encima. Ta mehanizem pojasnjuje hiter učinek hormonov, zlasti glukagona, na raven fruktozo-2,6-bisfosfata v celici (glej 16. poglavje).
Nekateri uravnavajo tudi aktivnost bifunkcionalnega encima
Priporočamo tudi
 Ioni so atomi, ki nosijo naboj. Pozitivno nabiti vodikovi ioni se imenujejo
Ioni so atomi, ki nosijo naboj. Pozitivno nabiti vodikovi ioni se imenujejo
 Kako ugotoviti natančen odmerek zdravila?
Kako ugotoviti natančen odmerek zdravila?
 Daniel najbolj škoduje otrokom
Daniel najbolj škoduje otrokom
-
Povzetek lekcije o literarnem branju na temo "Samostojno branje
 Vse naloge, naloge za igro, piškoti!
Vse naloge, naloge za igro, piškoti!
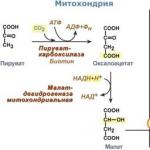 Sinteza glukoze iz aminokislin Glukoneogeneza iz glutaminske kisline
Sinteza glukoze iz aminokislin Glukoneogeneza iz glutaminske kisline